BioAcyl Corp |
|
Gusev, E., & Zhuravleva, Y. (2022). Inflammation: A New Look at an Old Problem. International Journal of Molecular Sciences, 23(9), 4596. Added by: Dr. Enrique Feoli (10/06/2022, 12:12) Last edited by: Dr. Enrique Feoli (09/01/2024, 18:31) |
| Resource type: Journal Article Published DOI: 10.3390/ijms23094596 ID no. (ISBN etc.): 1422-0067 BibTeX citation key: Gusev2022 View all bibliographic details |
Categories: BioAcyl Corp, BioAcyl Corp Subcategories: inflammation, Stress response Creators: Gusev, Zhuravleva Collection: International Journal of Molecular Sciences |
Views: 6/861
|
| Abstract |
|
Pro-inflammatory stress is inherent in any cells that are subject to damage or threat of damage. It is defined by a number of universal components, including oxidative stress, cellular response to DNA damage, unfolded protein response to mitochondrial and endoplasmic reticulum stress, changes in autophagy, inflammasome formation, non-coding RNA response, formation of an inducible network of signaling pathways, and epigenetic changes. The presence of an inducible receptor and secretory phenotype in many cells is the cause of tissue pro-inflammatory stress. The key phenomenon determining the occurrence of a classical inflammatory focus is the microvascular inflammatory response (exudation, leukocyte migration to the alteration zone). This same reaction at the systemic level leads to the development of life-critical systemic inflammation. From this standpoint, we can characterize the common mechanisms of pathologies that differ in their clinical appearance. The division of inflammation into alternative variants has deep evolutionary roots. Evolutionary aspects of inflammation are also described in the review. The aim of the review is to provide theoretical arguments for the need for an up-to-date theory of the relationship between key human pathological processes based on the integrative role of the molecular mechanisms of cellular and tissue pro-inflammatory stress.
|
| Notes | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
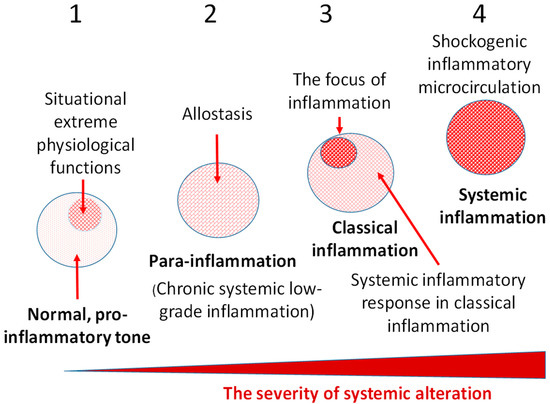
Variants of tissue pro-inflammatory stress. 1—Physiological variants of TS; 2—Non-classical low-grade inflammation (para-inflammation), which at systemic level may be manifest as stably altered homeostasis (allostasis); 3—Classical inflammation (the organism’s response to a significant local injury) is characterized by the presence of its attribute—a focus of inflammation and, in some cases, a systemic inflammatory response aimed at resourcing the focus of inflammation; 4—Life-critical systemic inflammation, the key phenomenon of which is a systemic microvascular response comparable in intensity to the local response in the focus of classical inflammation. Evolutionary Trends in the Development of InflammationUnderstanding of the evolutionary patterns in the emergence and development of mechanisms of inflammation and innate and adaptive immunity is important for the holistic characterization of inflammation as a general pathological process. The following stages may be distinguished in the development of the inflammatory process in the evolution of species:
These evolutionary differences seem to manifest themselves most clearly in the development of sepsis. Thus, systemic bacterial infections in fish and amphibians are characterized by microbial colonization of the gill, skin, muscles, internal organs, as well as their erosions, ulcerations, necroses, vascular damage, and hemorrhages [339,340,341]. Amphibians additionally show a more intense exudative reaction with fibrin accumulation in infected organs [342]. In reptiles, the generalization of infection is associated with multiple granulomas in internal organs, and with a large number of heterophils (mammalian neutrophil analogues) in granulomas during extracellular bacterial infections and predominantly lethal lesions of the heart and central nervous system [343,344]. In birds, generalized infection also presents as secondary microbial colonization; often affects the endocardium and myocardium; and reveals fibrinous deposits in tissues and granulocytic infiltrates, the major causes of death being thromboembolism in vital organs and septic endocarditis [343,345,346]. In humans, however, secondary microbial colonization of internal organs, and even bacteremia, is not a necessary condition for death in sepsis [347,348]. In fact, the association between bacteremia and endotoxemia is not always detected in patients [349]. In dogs, cats and various other mammalian species, the typical features of infectious and aseptic critical states are the accumulation of cytokines and other phlogogenic factors in the blood, systemic microthrombosis and microvascular activation during shock and multiple organ dysfunction, in some cases without signs of secondary pyemia [350].
Thus, the processes of para-inflammation, different variants of classical inflammation, and life-critical systemic inflammation have arisen at different stages of evolution. This evolutionary division is an additional argument for the need to comprehensively describe the general patterns of these processes, but also to differentiate these typical pathological processes in humans.
Note: yes—presence of a sign; no—absence of a sign; yes/no—sign detected in individual species; “?”—no reliable data on the phenomenon as a whole, but individual manifestations are possible; PRR—pattern recognition receptors; Ig—immunoglobulin; SIR—systemic inflammatory response; NES—neuroendocrine system; 1—e.g., parasite encapsulation [310]; 2—separation of lymph and blood; 3—the absence of a system of microcirculatory units; 4—in the most evolutionarily developed fish [333]; 5—only in mammals, complement anaphylatoxins (C3a and C5a) are formed in the liquid phase of the blood, for example, under the influence of hemostasis factors (XIIa, plasmin and thrombin) [334]; 6—only mammals have an extrinsic pathway for hemostasis activation (associated with the appearance of binding factor XI in them), and there are significantly fewer triggering factors (V, VII, and a soluble form of tissue factor) in plasma in birds than in mammals [335]; 7—in some invertebrates, some cytokine-like factors may be detected in hemolymph and other tissues, but there is no developed cytokine network; 8—DTH in birds is associated with the presence of high-affinity Fc receptors to IgY (FcυR) on mast cells [336], but DTH is significantly slower in birds than in mammals; 9—in this case, systemic inflammation is seen as a general pathological process with a systemic ‘inflammatory microcirculation’ phenomenon, not as a synonym for SIR; 10—according to the theory of G. Selye [337,338].
Added by: Dr. Enrique Feoli Last edited by: Dr. Enrique Feoli |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||